
The following is a copy of articles published by Cornell University.PIGEON NAVIGATIONThe ability of homing pigeons to navigate back to their "home" loft from distant and unfamiliar release sites has provided an excellent means by which to investigate the importance of various external cues in spatial navigation. When reared in a particular loft, a young pigeon can be transported hundreds of miles away and successfully find its way home from the release site. Because it requires the pigeon to pinpoint a specific location, this behavior necessitates more than the compass orientation system of migratory birds. Instead, the pigeon must be able to determine its position relative to the location of the home loft in order to orient itself in the proper direction. In doing so, pigeons use a variety of external cues such as the sun, visual landmarks, olfactory cues, and the earth's magnetic field. Depending on the weather conditions, where the pigeon was raised, and the nature of the release site, pigeons use a combination of these cues to determine their flight path. Examining when and how different cues are utilized by homing pigeons has provided neuroethologists with a better understanding of avian spatial navigation. HOMING BEHAVIORIntrinsically, pigeon homing behavior is well suited to neuroethological study. Because the natural environment of the homer is a domestic one, it is easier to observe and manipulate pigeons in their normal surroundings without drastically affecting their natural behavior and setting. As a result, homing pigeons have become a favorite subject for the study of avian spatial navigation. Many ingenious methods for investigating the use and salience of various external cues in pigeon homing have been developed. From these experiments, we have gained no definitive explanation for the ability of homers to navigate back to their home loft, but rather we have learned that the mechanism underlying this behavior is extremely complex. Many experiments have yielded different and often contradictory results which make it difficult to develop a universal understanding of the homing mechanism. It is clear, however, that pigeons are able to utilize several sources of environmental information in order to home efficiently under variable conditions. Of these cues, the most well studied are the pigeon's use of the sun, environmental odors, the magnetic field and visual landmarks. Gould (1982) describes the behavior of a homing pigeon as it is released from a site distant from its home loft. Initially, the pigeon circles around the release site several times. It then orients itself in a particular direction and flies in a relatively strait path away from the release site. The original bearing in which the pigeon flies is usually quite close to the actual direction of the home loft. Although the approximate direction is maintained during flight, the homeward journey of the pigeon varies. A pigeon, even when it is released from the same site on numerous occasions, does not fly an identical route each trip. The original flight direction is often referred to as the vanishing bearing or bearing at which the bird disappears from visual or radio contact. Because it is a reliable measure of the overall flight direction, the vanishing bearing is often used in the experimental literature to represent the homeward orientation. When many birds are released at a particular site, the mean vanishing bearing can be calculated by averaging the vanishing bearings of the individual pigeons. Interestingly, the mean vanishing bearing at a particular site often deviates slightly from the actual homeward direction. At each site, this deviation, known as release site bias, is reproducible and always occurs in the same direction (i.e. left or right) of the home loft bearing. In many instances, a mean vector length is calculated. The direction of the vector corresponds to the mean vanishing bearing and its length indicates the significance of the directional measure. If most pigeons are oriented in roughly the same direction, the mean vector length will be longer than when the vanishing bearings of the pigeons are highly variable. Most pigeon homing experiments manipulate the pigeons' ability to utilize one or more external cues and subsequently release the birds at a site distant from the home loft. Using the measures described above as well as others, experimenters can determine the effect, if any, of the experimental manipulations on homing behavior. Gould (1982) describes three theories that have been proposed to explain the navigational abilities of homing pigeons. Firstly, it may be that pigeons, like ants and honey bees, are able to track their outward journey so that they can later retrace their path home. Secondly, pigeons could home based on a cue gradient system which is centered at the home loft. Such a system, however, would not require a compass mechanism. Lastly, the pigeon may home via a true map sense. This would enable the pigeon to determine its location directly through site specific cues which would describe its position in relation to the home loft. Because pigeons deprived of outward journey information are able to home well, the first hypothesis has been largely disproved (Wallraff, 1980). Furthermore, pigeons have been shown to possess an internal compass mechanism which they use for spatial navigation (Kramer, 1950). It appears, then, that the last theory is most likely the correct one in the context of pigeon homing. In fact, the theory of true navigation in pigeons has been almost unanimously supported by the experimental literature. This type of navigation is often described as using a map and compass. Kramer (1953) proposed this two-step mechanism based on the theory that in order to find its way home, the pigeon must first be able to extrapolate its map position. From this determined map position, the pigeon can then orient itself in the home direction via some sort of compass mechanism. Although there is much controversy surrounding the various environmental stimuli that may be used by pigeons, the evidence indicates that the map component used for site localization is based on olfactory, magnetic, and visual cues while the compass sense appears to be guided by the sun and magnetic information (Wallraff 1990). VISUAL CUESThe importance of visual cues in pigeon homing has received little attention compared to that given olfactory and magnetic cues. Perhaps this is a result of some early experiments in which pigeons equipped with frosted contact lenses were found to home well and were able to locate their home lofts with excellent accuracy when denied access to visual information (Keeton, 1974). Given these results and the fact that pigeons are able to home from unfamiliar areas where there are no apparent familiar landmarks, experiments naturally focused their attention elsewhere. Recent experiments, however, have shown that visual landmarks are an important aspect of the pigeons' spatial map of the familiar area (Chappell and Guilford, 1997; Burt et al., 1997). Using an indoor, food searching task Chappell and Guilford investigated the pigeon's ability to use visual landmarks to locate food. They found that pigeons were indeed able to use visual landmarks in such a task, but only if these landmarks were three and not two dimensional. Because use of two dimensional visual cues all together prevented the pigeons from acquiring the task, it seemed that the pigeons were not able to use magnetic field information in this experiment. These results suggested that pigeons can use visual cues in a spatial learning task without access to the sun. The authors further suggest that their findings lend support to Wallraff's conclusion that anosmic pigeons are able to home from familiar sites due to their ability to use visual landmarks to navigate home (Chappell and Guilford, 1997). Burt et al. (1997) also found that pigeons use visual information when released from familiar sites distant from the home loft. This experiment allowed the experimental pigeons visual access to the release site for five minutes prior to the actual release. The homing speeds of these pigeons were then compared to those of the control group which was not allowed visual access to the site before release. It should be noted that both groups of birds were allowed access to olfactory information during the previewing period. The experimental birds homed, on average, sixteen percent faster than the controls indicating that their previewing experience allowed them to home more efficiently. Visual landmarks, then, seem to be a component of the pigeon's ability to discern the homeward direction at a familiar release site. The types of landmarks and the specific features of those landmarks that are used to form this visual map, however, are not known with any degree of certainty. MAGNETIC CUESWilliam Keeton (1969) noted that pigeons must be using another system in addition to the sun compass to determine directional information. On overcast days, when the sun is not visible, pigeons are able to home quite well. In fact, he found that clock shifted pigeons deviate from normal pigeons only on sunny days. During overcast, however, the orientation of the clock shifted and control pigeons were both homeward bound and there was no significant difference between the groups. Thus, he concluded that the sun is used to obtain directional information when it is visible, but that pigeons must have a secondary compass sense. What directional information might they be obtaining on cloudy days? Keeton hypothesized that pigeons may have the ability to detect the earth's magnetic field. In 1971, he attempted to disrupt this detection by gluing a bar magnet onto the pigeons. He found that these pigeons were often unable to navigate home from unfamiliar release sites under overcast conditions but experienced little difficulty on sunny days when compared to control birds that were equipped with a brass bar.h magnet. In five out of the six releases, the mean vanishing bearing of birds with magnets was either random or not homeward oriented while control birds were oriented well in all but one release. Thus, the magnets often interfered with the normal navigation mechanism of the pigeons. Keeton concluded that there seem to be two compass systems utilized during navigation with the primary system relying on the position of the sun and the secondary one on the earth's magnetic field. From these magnet experiments came an interesting finding regarding the development of the compass sense in pigeons. Keeton found that young, inexperienced pigeons were often disoriented even in sunny conditions as a result of the magnets. He, and later others, attributed this result to the theory that the magnetic compass is innate and used to calibrate the sun compass through homing experience. These first flight birds were therefore unable to use the sun compass to obtain directional information because of their inexperience and could not navigate via magnetic field detection because of the attached magnet (Gould, 1982; Keeton, 1974). Another famous set of experiments was conducted in which an artificial magnetic field was created around the pigeon's head using a Helmholtz coil (Walcott and Green, 1974). When this artificial field was oriented with the north pole pointing upwards, pigeons often flew 180 degrees away from home under overcast conditions, while their homing ability was unaffected under clear skies. The pigeons equipped with coils which had the magnetic south pole pointing upwards were able to home accurately on both overcast and sunny days. (See results below) 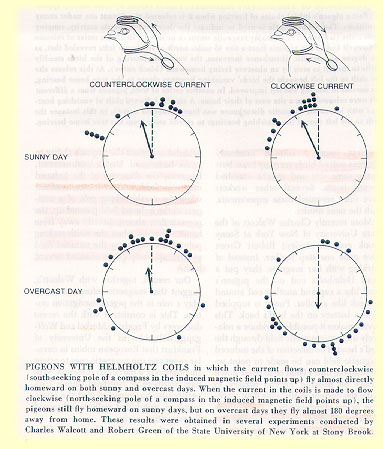 In a similar experiment, Walcott (1977) used Helmholtz coils to investigate the effect of applying an artificial magnetic field under clear skies. He found that a field of 0.1, 0.3, or 0.6 gauss created a detectable difference between control and experimental birds on sunny days. The length of the unit vector was decreased in both Nup and Sup pigeons when compared to the length of the unit vector of the control birds. Walcott concluded that this increase in scatter indicated a functional interaction between the magnetic and sun compasses. It appeared that the applied magnetic field was able to produce variability in the vanishing bearings of pigeons even when the sun was fully visible. Perhaps, then, the sun and magnetic compasses do not act completely autonomously, but rather they are functionally integrated by the pigeon. Gould (1982), Wiltschko (1991), and Walcott (1996) discuss the possibility that magnetic cues could be used by pigeons in the context of a map sense. Since the Earth's magnetic field varies in strength and dip angle over its surface, a pigeon able to detect small variation in either of these components could create a spatial map using this information. Keeton et al. discovered that pigeons released from the same site on different days vanished at slightly different bearings on each day. This effect was termed the "K-effect" and was not apparent when magnets had been fixed on the birds prior to release (Keeton, 1974). The degree to which the mean vanishing bearing changed from one day to another was correlated with the variation in the magnetic field before the release. Because the fluctuations in the magnetic field are so small from day to day and the K-effect appeared under sunny skies, it seems unlikely that the deflection of the vanishing bearings could be due to effects on the magnetic compass. Additionally, Gould argues that the exaggeration of release site biases during magnetic storms (natural magnetic disturbances that result from solar activity) show that pigeons are indeed sensitive to tiny changes in magnetic field properties which would be required for their use of magnetic information in forming a map. The theory of a magnetic map is also supported by the behavior of pigeons released at magnetic anomalies. When released at anomalous sites, pigeons reared in certain lofts will scatter randomly. It appears that the magnetic disturbance at these particular sites leaves the pigeons completely disoriented, thus it is likely to affect the map mechanism. Interestingly, groups of pigeons raised in separate loft locations experience magnetic anomalies differently. Walcott (1996) discusses his finding that pigeons raised at one loft were entirely disoriented when released at a particular anomaly while pigeons raised at a different loft nearby oriented well when released at the same site. He goes on to suggest that this difference may result from differences between lofts, rather than pigeons. It seems that pigeons reared in different lofts learn to use different external cues during navigation. This may be a function of the availability of types of information at the home loft. Pigeons that learn to rely on magnetic cues experience difficulty at magnetic anomalies while pigeons that learn to use some other cue do not. The magnetic map hypothesis has been refuted on several grounds. Firstly, most attempts to train pigeons to respond to a change in the magnetic field have failed (Couvillon et al., 1992; Kreithen, 1975). These experiments are based on the assumption that if a pigeon is able to detect variation in the magnetic field, then they should be able to be trained via classical conditioning to respond to such a change. In his Ph.D. thesis, Kreithen (1975) tested 97 pigeons for their ability to detect changes in the magnetic field using a classically conditioned increase in heart rate as the conditioned response. Here, pigeons were placed in a sealed container and first exposed to a change in the magnetic field (induced with Helmholtz coils) followed by a weak electric shock which caused the pigeons' heart rate to increase.
Despite these criticisms, much of the behavioral evidence implicates the use of the geomagnetic field in pigeon homing behavior. The search for the mechanism of detection of the magnetic field in birds has therefore been under investigation for some time. Although no specific magnetic sense organ has been identified, speculation that magnetic field detection may involve the retinal photorecteptors as well as a particle based magnetic field receptor has been described (Beason, 1995; Leask 1977). MAGNETIC FIELD DETECTIONSeveral experiments have focused on the anatomy underlying magnetic field detection in the pigeon. Investigation has centered around the photopigments of the visual system and magnetite particles associated with the trigeminal nerve. Before this type of research began, a theoretical physicochemical mechanism for magnetic field detection was proposed (Leask 1977). This theory is based on an optical pumping mechanism by which the retina can detect radiation in the visible spectrum. It was proposed that a molecule, such as the rhodopsin molecule, would have a magnetic moment in its triple state. The energy of these molecules, in their triple state, varies with the strength and direction of the magnetic field. A magnetic detection system may function by detecting the preferred field direction and intensity of the triple states of molecules within the visual system when the animal is oriented in different directions with respect to the magnetic field. Devising an experiment based on this theory, Semm and Demaine (1986) showed that neurons in the visual system responded to changes in the direction of the magnetic field. They proposed that these responses contained directional information because cells in the optic tectum and the nucleus of the basal optic root (nBOR) were selectively active according to the direction of the magnetic stimulation. Specific cells within both optic tectum and nBOR appeared to respond within a certain range of magnetic field direction when the direction of the field was gradually inverted. 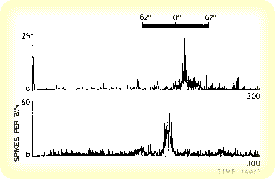 It has also been suggested that birds may use ferromagnetic compounds such as magnetite in order to detect changes in the magnetic field (Beason et al. 1994). An experiment was conducted in which Bobolinks were magnetized with a magnetic pulse. This treatment resulted in different orientations depending on the direction of the magnetic pulse that was applied. Treatment with a second pulse, opposite in direction to the first, resulted in random orientations of the birds. Because the orientation of the birds could be affected by magnetization with a single pulse, these results support the idea that bobolinks are using a particle based magnetoreception system for navigation. The authors propose that the magnetite particles associated with the trigeminal nerve may be responsible for this type of magnetic field detection. SUN COMPASSExperiments conducted over the past fifty years elucidate the importance of the sun as navigational cue in pigeon homing. Kramer (1950) showed that the sun was used by pigeons in experiment which manipulated the sun during a spatial learning task. He trained pigeons to located food in a particular direction using the sun as a directional cue. When he deflected the image of the sun with mirrors, the orientation of the birds was correspondingly shifted. Hoffmann then found that the sun is used in conjunction with the pigeon's own internal clock. He shifted the circadian rhythm of birds by exposing them to artificial light, a procedure known as clock-shifting. He found that the orientation of the birds shifted approximately fifteen degrees for every hour that the bird's clock had been shifted. This deviation corresponds to the difference between the actual angular position of the sun and the position predicted by the birds own internal clock. Based on this finding, Matthews (1953) proposed the sun-arc hypothesis of navigation which conflicted with Kramer's map and compass hypothesis. 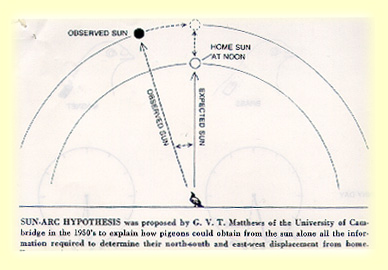  |